The simple act of eating involves a remarkably complex orchestra of physiological processes that extend far beyond the basic mechanics of chewing and swallowing. When you consume a meal, your body initiates an intricate cascade of neurological, hormonal, and mechanical responses designed to optimise digestion and regulate appetite. The speed at which you eat profoundly influences these delicate systems, potentially disrupting the natural rhythm your digestive tract has evolved to follow over millennia.
Modern eating habits, characterised by hurried consumption and minimal chewing, create a mismatch between our biological programming and contemporary lifestyle demands. This disconnect has significant implications for digestive health, metabolic function, and long-term wellbeing. Understanding how eating velocity affects the sophisticated communication networks within your body provides valuable insights into optimising nutritional outcomes and maintaining healthy weight regulation.
Neurophysiology of mastication rate and vagal nerve signalling
The relationship between chewing speed and digestive efficiency begins in the oral cavity, where mechanoreceptors embedded in the periodontal ligaments and jaw muscles detect the force and frequency of mastication. These specialised sensory organs play a crucial role in initiating the complex sequence of events that prepare your digestive system for incoming nutrients.
Mechanoreceptor activation in oral cavity during rapid versus slow chewing
When you chew rapidly, mechanoreceptors in the oral cavity receive compressed, intense stimulation that fails to trigger the full spectrum of preparatory digestive responses. Research indicates that optimal mechanoreceptor activation requires sustained, rhythmic pressure patterns that occur naturally during slower, more deliberate mastication. These receptors, particularly those located in the temporomandibular joint and surrounding musculature, respond differently to varying chewing velocities, influencing downstream digestive processes.
The duration and intensity of mechanoreceptor stimulation directly correlates with the magnitude of vagal nerve activation. Prolonged chewing stimulates these receptors more comprehensively, leading to enhanced parasympathetic nervous system engagement and improved digestive preparedness.
Vagus nerve transmission pathways and gastric motility modulation
The vagus nerve serves as the primary communication highway between the brain and the digestive organs, transmitting signals that coordinate gastric acid secretion, enzyme release, and peristaltic movements. When eating occurs too rapidly, this critical neural pathway experiences disrupted signalling patterns that can compromise digestive efficiency.
Vagal tone, the strength and consistency of vagus nerve signalling, becomes impaired when mechanoreceptor stimulation is insufficient due to rapid consumption. This impairment manifests as reduced gastric acid production, delayed enzyme activation, and suboptimal coordination of digestive organ function. The consequences extend beyond immediate digestive discomfort to include long-term metabolic implications.
Hypothalamic satiety centre response to trigeminal sensory input
The trigeminal nerve, responsible for facial sensation and mastication control, provides essential sensory feedback to the hypothalamic satiety centres. This feedback mechanism helps regulate appetite and food intake by communicating information about food texture, volume, and oral processing time to the brain’s appetite control regions.
Rapid eating significantly reduces trigeminal sensory input duration, leading to inadequate stimulation of hypothalamic satiety centres. Research demonstrates that individuals who consume meals quickly show delayed activation of these brain regions, resulting in continued appetite stimulation even after consuming adequate calories. This neurological delay contributes to overeating and poor appetite regulation.
Parasympathetic nervous system regulation through cephalic phase responses
The cephalic phase of digestion encompasses all preparatory digestive responses that occur before food reaches the stomach, including saliva production, gastric acid secretion, and pancreatic enzyme release. These responses depend heavily on adequate parasympathetic nervous system activation, which requires sufficient time and appropriate stimulation patterns.
Cephalic phase responses become markedly diminished when eating velocity exceeds the nervous system’s capacity to fully engage these preparatory mechanisms. The parasympathetic nervous system requires approximately 15-20 minutes to achieve optimal activation levels,
which is one reason why nutrition researchers so often emphasise the importance of taking at least 20 minutes to eat a meal. When you rush through a plate of food in five to ten minutes, the cephalic phase is cut short, salivary and gastric secretions are blunted, and pancreatic output lags behind the arrival of nutrients in the small intestine. Over time, this pattern can contribute to symptoms such as bloating, reflux, and erratic blood glucose excursions after meals. By contrast, slower eating supports a more robust parasympathetic response, smoother digestive transit, and more timely activation of satiety signals.
Gastric emptying kinetics and antral contractions in fast eaters
Once food passes through the oesophagus, eating speed begins to influence the mechanical behaviour of the stomach. Gastric emptying kinetics—the rate at which food leaves the stomach and enters the duodenum—depend on how efficiently the stomach can receive, store, and grind incoming material. Rapid eating challenges these processes by delivering large boluses of poorly chewed food in a short period, forcing the stomach to adapt under time pressure. This can disrupt the normal coordination between fundal accommodation, antral grinding, and pyloric control that underpins healthy digestion.
Fundal accommodation impairment due to rapid food bolus entry
The fundus, the upper reservoir portion of the stomach, is designed to relax and expand in a controlled way as food arrives, a process known as gastric accommodation. When you eat slowly, this relaxation is gradual, allowing intragastric pressure to remain relatively low while still accommodating meal volume. In fast eaters, however, food boluses arrive in quick succession, and the fundus may not fully relax before the next load arrives. This leads to transient increases in gastric pressure that can be perceived as early fullness, heaviness, or discomfort.
Impaired fundal accommodation also alters how evenly food is distributed within the stomach. Instead of forming a well-layered mixture that can be progressively processed by antral contractions, large clumps tend to sink and overload the distal stomach. Clinical studies using barostat techniques have shown that individuals with reduced fundal compliance experience more postprandial symptoms and are more prone to functional dyspepsia. By simply slowing down and allowing time between bites, you give the fundus a better chance to relax appropriately and buffer meal volume without excessive pressure spikes.
Pyloric sphincter dysfunction and duodenal reflux mechanisms
The pyloric sphincter regulates the passage of chyme from the stomach into the duodenum, opening and closing in synchrony with antral contractions and duodenal feedback signals. In conditions of rapid eating and incomplete trituration of food, the pylorus is confronted with larger, less homogeneous particles and higher intragastric pressure. To protect the small intestine from overload, the pylorus may respond with more frequent, rapid closures, leading to a “stop–start” emptying pattern that feels like delayed digestion.
In some individuals, this disrupted pattern can predispose to duodenogastric reflux, where bile and pancreatic secretions flow backwards into the stomach. This reflux not only aggravates heartburn and nausea but can also further impair gastric motility via inflammatory and sensory pathways. Observational data suggest that people who habitually eat very quickly report higher rates of reflux-like symptoms, and pyloric dysregulation is one proposed mechanism. A slower eating pace, combined with thorough chewing, produces a finer, more cohesive chyme that the pylorus can manage more efficiently, reducing mechanical strain on this critical gateway.
Gastric acid secretion patterns in tachyphagic eating behaviours
Gastric acid secretion is tightly regulated by neural, hormonal, and local factors that respond to meal composition and timing. During slow, mindful eating, acid production ramps up in a phased manner, matching the gradual arrival of food in the stomach. In tachyphagic (very fast) eating behaviours, the cephalic phase—driven by vagal signalling—is shortened, while the gastric phase is abruptly stimulated by a sudden flood of nutrients and distension. This mismatch can result in an initial underproduction of acid followed by compensatory hypersecretion.
Such irregular patterns may contribute to both poor protein digestion and increased risk of acid-related symptoms. When large volumes of food arrive before adequate acid is present, early digestion is inefficient, prolonging gastric residence time. As acid production catches up, intragastric pH may drop sharply at a point when the stomach is already distended, exacerbating discomfort and reflux susceptibility. Consistent evidence links rapid ingestion with higher rates of dyspepsia and heartburn, while controlled trials have shown that slowing eating speed can improve subjective digestive comfort even without changing what is eaten.
Antral wave propagation velocity changes during accelerated consumption
The antrum, the lower muscular portion of the stomach, generates powerful peristaltic waves that grind solid food into fine particles suitable for passage through the pylorus. Under normal conditions, these waves follow a predictable frequency and velocity, ensuring efficient mechanical digestion. However, when eating speed is high and large undigested boluses accumulate rapidly, antral contractions may adapt by increasing amplitude and altering propagation velocity to cope with the excess load.
High-resolution manometry studies suggest that in such scenarios, antral waves can become more forceful yet less coordinated, much like a conveyor belt that suddenly speeds up and starts to jostle its cargo. This can lead to pockets of poorly processed food coexisting with over-ground material, disrupting the orderly progression of chyme. You may perceive this as “food sitting in the stomach” despite having finished eating some time ago. By extending meal duration and allowing more gradual filling, you support more regular antral contractile patterns, better mixing of gastric contents, and a more predictable gastric emptying curve.
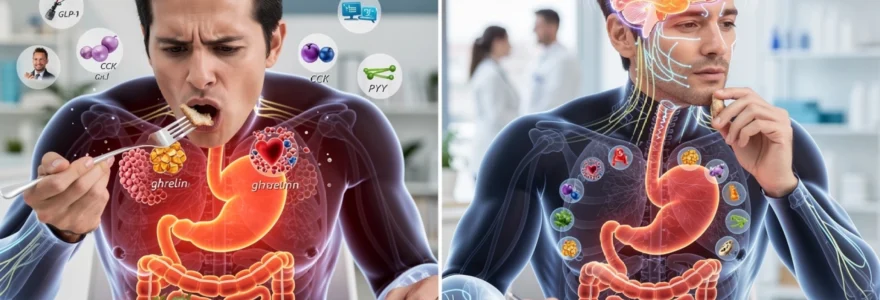
Hormonal cascade disruption: GLP-1, CCK, and ghrelin response timing
Beyond mechanical digestion, meal timing and eating speed profoundly affect the hormonal milieu that governs hunger, fullness, and metabolic regulation. Satiety hormones such as cholecystokinin (CCK), glucagon-like peptide-1 (GLP-1), and peptide YY (PYY) rise in response to nutrient exposure in the gut, while ghrelin—a key hunger hormone—falls as the stomach distends and energy is sensed. Because these hormones operate on a timescale of minutes to tens of minutes, rushing through a meal can create a temporal mismatch between energy intake and hormonal feedback.
In practical terms, this means you can easily consume more energy than your body actually needs before your hormonal “brakes” have time to engage. Several controlled feeding studies have shown that simply extending meal duration from around 10 minutes to 20–30 minutes increases postprandial CCK, GLP-1, and PYY levels and reduces subsequent energy intake. Understanding these hormonal response timings helps explain why eating slowly is such a powerful, low-tech tool for appetite regulation and weight management.
Cholecystokinin release latency and small intestinal l-cell activation
Cholecystokinin is released primarily from I-cells in the duodenum and jejunum when fats and partially digested proteins arrive. It promotes gallbladder contraction, pancreatic enzyme secretion, and notably, signals satiety to the brain via vagal afferents. There is an inherent latency between the start of a meal and peak CCK release, typically on the order of 15–30 minutes, because nutrients must transit from the stomach into the upper small intestine at a controlled rate.
When you eat rapidly, large quantities of food enter the stomach before gastric emptying has had time to deliver nutrients steadily to I-cells. As a result, the CCK signal that would naturally slow eating and enhance satiety lags behind actual intake. Some imaging and hormone studies suggest that slow eaters achieve higher CCK concentrations for the same meal, likely because prolonged mastication and more gradual gastric emptying increase the effective exposure of nutrients to I-cells. From a behavioural standpoint, spacing bites, chewing thoroughly, and pausing during a meal are all strategies that help align CCK release with real-time intake, reducing the risk of overshooting comfortable fullness.
Glucagon-like peptide-1 secretion delays in rapid consumption patterns
GLP-1, secreted by L-cells predominantly in the distal small intestine, acts as an incretin hormone enhancing insulin secretion, slowing gastric emptying, and promoting satiety. Because many GLP-1–producing cells are located further along the gut, there is an even longer delay between starting a meal and achieving peak GLP-1 levels. In fast consumption patterns, most of the meal may already have been eaten by the time GLP-1 begins to exert its appetite-suppressing and glucose-modulating effects.
Randomised trials comparing slow versus rapid eating have demonstrated higher GLP-1 responses and better postprandial glycaemic control when meals are consumed over 20–30 minutes. For individuals with impaired glucose tolerance or early type 2 diabetes, this timing difference can be clinically meaningful. Slowing eating speed is not a substitute for medication where required, but it can enhance endogenous GLP-1 signalling in a way that complements other metabolic therapies. Seen this way, your fork and chewing pattern become a low-cost “delivery system” for optimising your own gut hormone responses.
Ghrelin suppression mechanisms and stomach distension sensitivity
Ghrelin, often dubbed the “hunger hormone”, rises before meals and falls as the stomach fills and nutrients are absorbed. The rate at which ghrelin is suppressed depends on both mechanical distension of the gastric wall and downstream nutrient sensing. In slow eating patterns, stomach expansion and early nutrient delivery to the intestine occur in a gradual, coordinated fashion, producing a steady, sustained decline in ghrelin that aligns with emerging satiety.
In contrast, rapid ingestion creates abrupt distension before appropriate hormonal and neural feedback systems are fully engaged. Paradoxically, some studies have observed that ghrelin suppression can be less complete after very fast meals, possibly because mechanoreceptors and chemoreceptors are overstimulated in a short window, then adapt. From your perspective, this can feel like lingering hunger or a desire to snack again soon after a large, quickly eaten meal. Training yourself to extend meal duration helps smooth ghrelin’s fall and stabilise hunger signals between meals.
Peptide YY production kinetics and ileal brake response timing
Peptide YY is another satiety hormone secreted from L-cells, particularly in the ileum and colon, in response to fat and protein reaching the distal gut. It is central to the so-called ileal brake—a feedback mechanism that slows gastric emptying and small intestinal transit when sufficient energy has been ingested. Because PYY secretion depends on nutrients travelling far down the intestinal tract, it is inherently delayed relative to meal onset.
With rapid eating and accelerated delivery of a large meal to the upper gut, gastric emptying may initially proceed more quickly, outpacing the development of the ileal brake. Only later, when sizable amounts of nutrients finally reach the ileum, does PYY surge and slow things down—by which time overeating may already have occurred. Slow, evenly paced meals support a more proportional PYY response, aligning this powerful brake with actual intake and contributing to more enduring satiety after eating. For people struggling with constant post-meal nibbling, paying attention not just to what they eat but to the tempo of eating can meaningfully influence PYY dynamics.
Enzymatic activity modulation through mastication duration
Mechanical breakdown of food in the mouth is the first step in digestion, and its quality shapes downstream enzymatic activity throughout the gastrointestinal tract. Salivary amylase begins starch digestion during chewing, while lingual lipase initiates fat breakdown. When chewing is brief and superficial, the surface area of food particles is reduced less effectively, and saliva has limited contact time with substrates. This means that by the time food reaches the stomach and small intestine, digestive enzymes must work harder and longer to achieve complete breakdown.
Extended mastication, by contrast, increases particle fragmentation and thoroughly mixes food with saliva, effectively “pre-processing” the meal. Experimental work has shown that chewing starchy foods more times (for example, 30–40 chews per bite instead of 10–15) enhances early amylase activity and results in smoother, more consistent rises in blood glucose rather than sharp spikes. Proteins and fats also benefit from this prolonged oral phase, as better mechanical disruption exposes more binding sites to pepsin, gastric lipase, and later pancreatic enzymes.
From a practical perspective, you can think of chewing as your personal, built-in blender and pre-digestion system. The more work you ask your teeth and jaw to do, the less burden you place on your stomach and pancreas. People with marginal pancreatic function, subclinical hypochlorhydria, or age-related declines in digestive capacity may find that a simple focus on chewing each bite more thoroughly significantly improves tolerance of meals. This is one reason why clinicians often recommend “eat slowly and chew well” as a frontline strategy for managing functional digestive complaints.
Clinical evidence from bariatric surgery outcomes and mindful eating interventions
While mechanistic research helps us understand how eating speed affects digestion and satiety signals at a physiological level, clinical data show how these mechanisms translate into real-world outcomes. Two areas provide particularly compelling evidence: bariatric surgery follow-up and structured mindful eating interventions. In both contexts, modifying eating behaviour—especially eating rate—has been associated with improved weight control, better glycaemic responses, and enhanced quality of life.
Interestingly, many of the behavioural rules given to bariatric surgery patients (such as taking small bites, chewing thoroughly, and extending meal time) echo traditional advice around slow eating, but with amplified consequences if not followed. Mindful eating programmes, on the other hand, show that even without surgery or medications, training people to pay attention to the pace and experience of eating can meaningfully shift appetite patterns and energy intake. Taken together, these clinical models highlight that how you eat is not a trivial detail but a modifiable therapeutic target.
Therapeutic applications of controlled eating velocity in metabolic disorders
Given the interplay between eating speed, digestion, and hormone signalling, it is not surprising that controlled eating velocity is emerging as a low-cost, accessible adjunct in managing metabolic disorders. Conditions such as obesity, type 2 diabetes, non-alcoholic fatty liver disease, and metabolic syndrome are all influenced by chronic patterns of energy intake and postprandial physiology. Slowing down eating offers a behavioural lever to improve satiety, moderate glycaemic excursions, and reduce overall caloric load without strict food restriction.
For individuals with insulin resistance, for example, extending meal duration can enhance endogenous incretin responses (like GLP-1), modestly slow gastric emptying, and blunt post-meal glucose peaks. Combined with balanced macronutrient composition and adequate fibre, this supports more stable blood sugar profiles. In obesity treatment, coaching patients to recognise early fullness cues and to pause between bites helps prevent passive overeating driven by delayed hormonal feedback. Some structured weight-management programmes now explicitly track eating rate as a habit variable alongside food choices and physical activity.
Implementing controlled eating velocity in practice does not require complex technology. Clinically, we might suggest simple cues such as putting down utensils between bites, taking a sip of water after every few mouthfuls, or using a timer to ensure that a main meal lasts at least 20 minutes. Wearable devices and smartphone apps that prompt mindful pauses or record bite rate are being trialled as digital health tools to reinforce these behaviours. Ultimately, by aligning the speed of eating with the natural timing of digestive and satiety signals, you create conditions in which your biology can work with you—rather than against you—in pursuit of better metabolic health.
 Good health cannot be bought, but rather is an asset that you must create and then maintain on a daily basis.
Good health cannot be bought, but rather is an asset that you must create and then maintain on a daily basis.
